Princip flushingu je jednoduchý: krátkodobý vyšší příjem energie a živin má stimulovat ovulaci, a tím zvýšit počet selat ve vrhu. Ačkoliv se metoda používá desítky let, moderní genotypy a intenzivní chovy přinášejí nové otázky – především kdy flushing nasadit a jak dlouho jej udržovat, aby byl maximálné účinný a zároveň neškodil.

Jak flushing působí
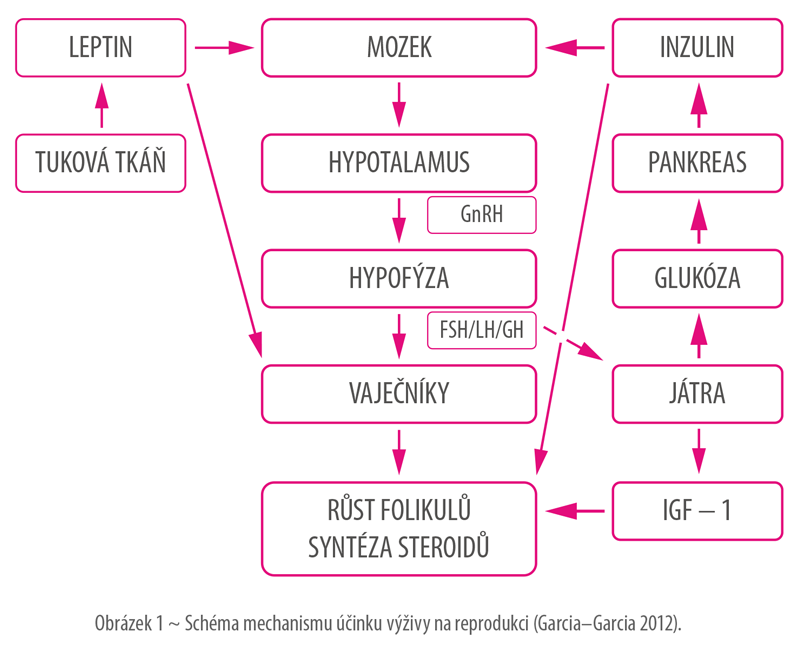
Zvýšený příjem energie v období před zapuštěním vede k (Van den Brand et al. 2000; Scaramuzzi et al. 2006; Zhou et al. 2010; Garcia–Garcia 2012; Langendijk & Peltoneimi 2019; Niblett 2024):
Důsledkem působení těchto mechanismů dochází ke zvýšení počtu všech narozených selat, kvality a vyrovanosti vrhu (Van den Brand et al. 2009) a kratšímu intervalu mezi odstavem a říjí (Tauson 2012). Účinek však není univerzální – reakce zvířat závisí na jejich věku, fázi reprodukčního cyklu a kondici. Prasničky a prasnice v dobré kondici reagují výraznějším nárůstem počtu ovulovaných vajíček. U zvířat ve vysoké kondici se může vyskytovat hyperglykémie, hypertenze, inzulinová rezistence a dyslipidémie, vedoucí k reprodukčním problémům (Newell–Fugate et al. 2015).
Prasnice
Flushing u prasnic je založený na obnově přirozeného reprodukčního potenciálu, který je vlivem laktace snížený. Během laktace jsou živiny, které prasnice přijímá, stejně jako její tělesné zásoby, využívány k produkci mléka. Jedná se o období s vysokým energetickým výdejem, a proto se zvíře během ní nachází v negativní energetické bilanci. V důsledku nedostatku energie dochází u prasnic k poklesu hadiny inzulinu, glukózy a leptinu, což následně vede k narušení folikulogeneze. Tyto změny alterují intrafolikulární tekutinu a snižují preovulační funkci a růst folikulů, což následně má vliv na kvalitu a počet oocytů (Tauson 2012).
Prasničky
Prasničky také představují výzvu, protože mladá zvířata musí vynaložit velké množství energie na růst. Z hlediska ekonomického, ale také k podpoření produktivity a dlouhověkosti je důležité, aby prasničky byly zapuštěny včas a v pozitivní energetické bilanci (Faccin et al. 2022). Pro včasné vyrovnání negativní energetické bilance v důsledku růstu je vhodné stejně jako u prasnic využít efektu flushingu. Moderní data potvrzují, že flushing má největší přínos právě u mladých prasniček v dobré kondici, tzn. ve věku mezi 230–240 dny, s hmotností mezi 140–160 kg a výškou hřbetního tuku mezi 13–15 mm (Theil et al. 2022). Právě u takových zvířat dochází při obohacení krmné dávky k největšímu nárůstu počtu ovulovaných vajíček (Garcia–Garcia 2012).
Důležitost vitamínů a minerálů
Reprodukční vlastnosti se dají podpořit i jinak než pouze zvýšením obsahu energie v krmné dávce, a to doplněním minerálních látek a vitamínů, které jsou nezbytné pro správné fungování metabolismu. K podpoření plodnosti patří z řad minerálů především zinek, hořčík, vápník, jód, selen, železo, měď a mangan. Komplexní interakcí těchto minerálů je regulována hormonální tvorba, funkce vaječníků, ovulace, oxidační stres, a implantace embrya do endometria (Kapper et al. 2024).
Mezi důležité vitamíny se řadí vitamín A a komplex vitamínů B. Vitamíny B2, B9 a B12 jsou klíčové pro správný vývoj a přežívání embryí, podporují syntézu proteinů a hormonální regulaci. Nedostatek jednoho z nich vede k hromadění homocysteinu, který embrya poškozuje.
Shastak & Pelletier (2024) ve své rešerši uvádí příklady se zvýšenou suplementací riboflavinu (B2) a jeho vlivu na reporodukční vlastnosti. Například Bazer (1990) ve své studii porovnával skupinu se suplemetovaným riboflavinem 100 mg/den v rané březosti (4–10 den) oproti skupině kontrolní bez suplementace. Výsledkem byla vyšší míra březosti, vyšší počet žlutých tělísek, vyšší počet všech a živě narozených selat. Následně zde byl i statisticky významný vliv na přežitelnost selat a hmotnost vrhu za 21 a 42 dní po porodu.
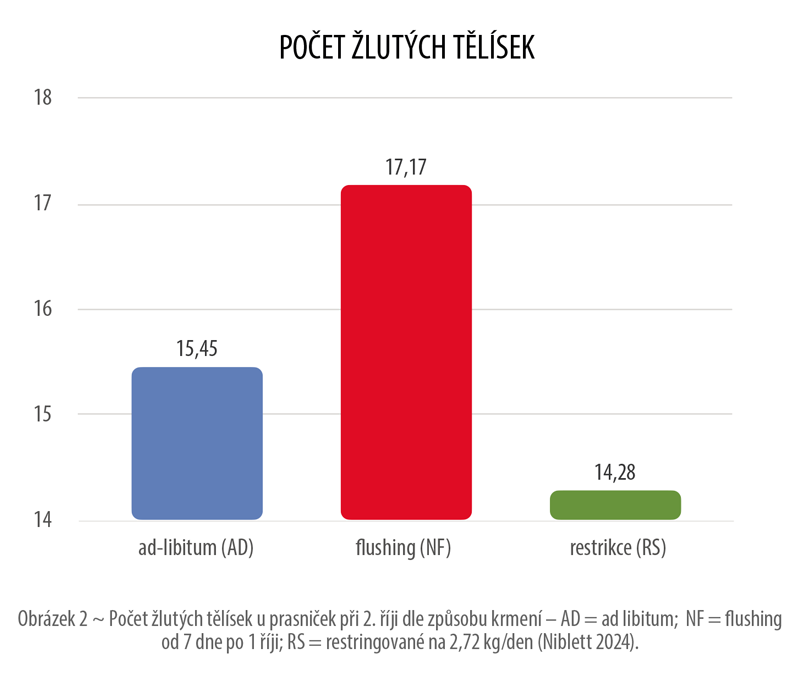
Při suplementaci vitamínu B9 bylo v několika doložených studiích pozorováno zlepšení reprodukčních ukazatelů prasnic: při dávce 1 ppm vzrostl počet narozených selat na prasnici (Lindemann a Kornegay 1989), zatímco při dávce 15 ppm došlo ke snížení embryonální úmrtnosti a ke zvýšení hladin prostaglandinu E2 (PGE2) a transformujícího růstového faktoru β2 (TGFβ2), mediátory významné pro ebryonální vývoj (Giguère et al. 2000; Guay et al. 2004).
Podle studie Simarda et al. (2004), která zkoumala vliv vitamínu B12 na velikost vrhu prasnic, bylo krmivo obohaceno o 20, 100, 200 a 400 ppb tohoto vitamínu. Výsledky ukázaly, že prasnice s příjmem vyšším než 100 ppb měly vícečetnější vrhy.

Vitamín A u prasniček podporuje vývoj oocytů urychlením meiózy a zvýšením podílu oocytů ve vyspělějších fázích. Whaley et al. (2000) ve své studii zkoumal vliv suplementace vitamínu A při dávkách 1x106 IU na vývoj oocytů, folikulární hormony a ovulaci. Výsledkem byly vyšší hodnoty u hormonů progesteronu, IGF–I, PGF2 a vyšší počet oocytů v pokročilejších vývojových stádiích. Tyto účinky mohou nasvědčovat zvýšené přežitelnosti embryí a větší velikosti vrhu.
Případová studie
Chen et al. (2012) ve své studii zkoumali mimo jiné vliv obohacení krmné směsi o cukry, tuky a Regumate na reprodukční vlastnosti prasnic a vlastnosti jejich následujícího vrhu. Mezi zkoumané parametry reprodukčních vlastností patřily intervaly odstav–říje a odstav–ovulace, doba trvání říje, míra březosti, velikost folikulů, obsah hormonů leptinu, estradiolu a progesteronu. Do vlastností vrhu byl zahrnut počet všech narozených selat, počet živě narozených selat a hmotnost selete při narození. Prasnice po prvním vrhu byly
devět dní před odstavem rozděleny na čtyři pokusné skupiny a to:
» skupina C – kontrola (n = 24),
» skupina F – 1 kg krmné směsi pro kojící prasnice nahrazen krmnou směsí bohatou na tuky (n = 23),
» skupina S – 1 kg krmné směsi pro kojící prasnice nahrazen krmnou směsí bohatou na cukry (n = 23),
» skupina R – přídavek Regumate, syntetického steroidního hormonu určeného k synchronizaci říje (n = 22),
U reprodukčních vlastností prasnic skupina S zaznamenala největší zkrácení intervalu odstav–říje (4,3 ± 0,1 dny) oproti skupině R (5,2 ± 0,1 dny; p < 0,05) a mírné zkrácení oproti skupině F (4,4 ± 0,1 dny; p > 0,05) a C (4,6 ± 0,3 dny; p > 0,05). Interval od odstavu do ovulace byl u skupiny S (5,7 ± 0,1 dne) statisticky významně kratší než u skupiny R (7,0 ± 0,1 dny; p < 0,05) a kratší také oproti skupinám F (6,2 ± 0,1 dny; p = 0,08) a C (6,2 ± 0,2 dny; p = 0,06). Skupina S zároveň vykazovala nejkratší dobu trvání říje (49,5 ± 2,4 hod.), avšak statisticky významný rozdíl byl zaznamenán pouze vůči skupině R (61,6 ± 2,5 hodin; p = 0,01). Velikost folikulů se významně nelišila mezi jednotlivými skupinami a při ovulaci dosahovala průměru 7 ± 0,1 mm (p > 0,05) u všech zvířat. Hladina leptinu byla u skupiny S druhá nejvyšší (3,06 ± 0,24 ng/ml), přičemž nejvyšší hodnoty byly naměřeny u skupiny R (4,67 ± 0,39 ng/ml), která se významně lišila od ostatních skupim (p < 0,05). Koncentrace estradiolu byla naopak u skupiny S nejnižší (6,62 ± 0,52 pg/ml), ovšem bez statisticky významného rozdílu (p > 0,05). Čtyři dny po ovulaci byla ve skupině S zjištěna nejvyšší hladina progesteronu (17 ± 0,1 ng/ml) ve srovnání s ostatními skupinami (p < 0,05). Mezi skupinami nebyl nalezen statisticky významný rozdíl u míry březosti, kdy skupina S měla březích zvířat 96 % a ostatní skupiny se pohybovaly 78–82 % březích zvířat (p > 0,05). Celkem 17 prasnic neudrželo březost, z toho sedm ve skupině C, pět ve skupině F, tři ve skupině R a dvě ve skupině S. Vliv na vlastnosti vrhu byl statisticky významný pouze ve srovnání v počtu všech narozených selat, který byl nejvyšší u skupiny S (12,4 ± 0,5) a nejnižší u skupiny C (10,7 ± 0,6; p = 0,04). Ve zbylých parametrech vrhu nebyl nalezen statisticky významný rozdíl, ovšem skupina S měla nejvyšší počet živě rozených selat (11,8 ± 0,5). Skupina S navíc vykazovala nejvyšší podíl vrhů s více než 10 selaty, a to 89 %. Doplnění krmiva cukry pozitivně ovlivnila reprodukční parametry prasnic. Skupina S vykázala nejkratší interval od odstavu k říji a ovulaci a vyšší hladiny progesteronu po ovulaci, což svědčí o rychlejším nástupu reprodukční činnosti. I když celková březost nebyla statisticky odlišná, skupina S měla vyšší míru březosti a oprašenosti, vyšší průměrný počet narozených selat a největší podíl vrhů s více než 10 selaty. Autor tyto efekty přisuzuje zvýšené hladině glukózy a inzulinu, které mohou přímo ovlivňovat folikulární vývoj a kvalitu oocytů, a tím zvyšovat početnost vrhu.

Závěr
Moderní poznatky ukazují, že obohacení krmné dávky o energii před zapuštěním je stejně tak důležité jako samotná výživa během laktace. Flushing je silný nástroj pro zlepšení reprodukce, ale jeho účinnost závisí na správném načasování, kondici zvířete a vyváženosti celé krmné dávky. Moderní genetika prasnic vyžaduje nejen zvýšení krmné dávky, ale i cílené dodání rychle dostupné energie a klíčových živin. Jedním z osvědčených řešení je náš Pig Flush – doplňková krmná směs pro prasničky a prasnice, která se podává:
» prasničky: pět dní před očekávanou říjí v dávce 200–250 g/ks/den
» prasnice: den po odstavu do zapuštění v dávce 200–250 g/ks/den
Hlavní benefity Pig Flush:
» zvýšení hladiny glukózy v krvi a tím i podpoření tvorby ostatních hormonů,
» rychlejší a výraznější říje – zkrácení inseminačního intervalu,
» rychlejší růst folikulů a zvýšený počet ovulovaných vajíček – početnější a vyrovnanější vrh.
Díky kombinaci dextrózy jako koncentrovaného zdroje pohotové energie, kvalitního oleje, vitamínů a organicky vázaných mikroprvků s vysokou vstřebatelností, přináší Pig Flush prasnicím a prasničkám přesně to, co potřebují v rozhodujícím období před zapuštěním. ≈
Zdroje:
AGWEEK. Online. 2023. Dostupné z: https://www.agweek.com/news/policy/supreme–court–decision–puts–pig–policy–under–california–control–but–the–fight–isnt–over. [cit. 2025–09–13].
Bazer FW. 1990. Use of supplemental dietary riboflavin to increase fertility and/ or prolificacy in animals. US patent US5063226A. IFI CLAIMS Patent Services.
Chen TY, Stott P, Bouwman EG, Langendijk P. 2012. Effects of Pre‐Weaning Energy Substitutions on Post‐Weaning Follicle Development, Steroid Hormones and Subsequent Litter Size in Primiparous Sows. Online. Reproduction in Domestic Animals 48:512–519.
Faccin JEG et al. 2022. Gilt development to improve offspring performance and survivability. Online. Journal of Animal Science 100:1–10.
Garcia–Garcia RM. 2012. Integrative Control of Energy Balance and Reproduction in Females. ISRN Veterinary Science 1–13.
Giguère A, Girard CL, Lambert RD, Laforest JP, Matte JJ. 2000. Reproductive performance and uterine prostaglandin secretion in gilts conditioned with dead semen and receiving dietary supplements of folic acid. Canadian Journal of Animal Science 80:467–472.
Guay F, Matte JJ, Girard CL, Palin MF, Giguère A, Laforest JP. 2004. Effect of folic acid plus glycine supplement on uterine prostaglandin and endometrial granulocyte–macrophage colony–stimulating factor expression during early pregnancy in pigs. Theriogenology 3:485–498.
Kapper C, Oppelt P, Ganhör C, Gyunesh AA, Arbeithuber B, Stelzl P, Rezk–Füreder M. 2024. Minerals and the Menstrual Cycle: Impacts on Ovulation and Endometrial Health. Nutrients 16:1008.
Langendijk P & Peltoneimi O. 2019. How does nutrition influence luteal function and early embryo survival. Bioscientifica Proceedings 68:145–158.
Lindemann MD, Kornegay ET. 1989. Folic acid supplementation to diets of gestating–lactating swine over multiple parities. J Anim Sci. 2:459–464
Newell–Fugate AE et al. 2015. Effects of Obesity and Metabolic Syndrome on Steroidogenesis and Folliculogenesis in the Female Ossabaw Mini–Pig. PLoS ONE 10:6.
Niblett RT. 2024. Feeding Strategies to Enhance Gilt Reproduction and Subsequent Longevity and Productivity in the Breeding Herd. [Dissertation]. Suffolk, Virginia Polytechnic Institute and State University.
Scaramuzzi RJ et al. 2006. A review of the effects of supplementary nutrition in the ewe on the concentrations of reproductive and metabolic hormones and the mechanisms that regulate folliculogenesis and ovulation rate. Reproduction Nutrition Development 4:339–354.
Shastak Y, Pelletier W. 2024. Exploring the role of riboflavin in swine well–being: a literature review. Porc Health Manag 10:46.
Simard F, Guay F, Girard C, Giguère A, Laforest JP, Matte JJ. 2004. La vitamine B12 chez la truie gravide: fautil actualiser le besoin? Journées Rech. Porcine en France 36: 229–234.
Tauson A. 2012. Reproduction. In: Nutritional Physiology of Pigs. s.l.:Videncenter for Svineproduktion, Landburg and Fodevarer.
Theil PK, Krogh U, Bruun TS, Feyera T. 2022. Feeding the modern sow to sustain high productivity. Molecular Reproduction and Development 90:517–532.
Van den Brand H, Dieleman SJ, Soede NM, Kemp B. 2000. Dietary energy source at two feeding levels during lactation of primiparous sows: I. Effects on glucose, insulin, and luteinizing hormone and on follicle development, weaning–to–estrus interval, and ovulation rate. Journal of Animal Science 78:396–405.
Van den Brand H, Van Enckevort L, Van Hoeven E, Kemp B. 2009. Effect of Dextrose Plus Lactose in the Sows Diet on Subsequent Reproductive Performance and within Litter Birth Weight Variation. Reprod. Dom. Anim. 4:884–888.
Whaley SL, Hedgpeth VS, Farin CE, Martus NS, Jayes FC, Britt JH. 2000. Influence of vitamin A injection before mating on oocyte development, follicular hormones, and ovulation in gilts fed high–energy diets. Journal of Animal Science 78:1598.
Zhou DS et al. 2010. Dietary energy source and feeding levels during the rearing period affect ovarian follicular development and oocyte maturation in gilts. Theriogenology 74:202–21.